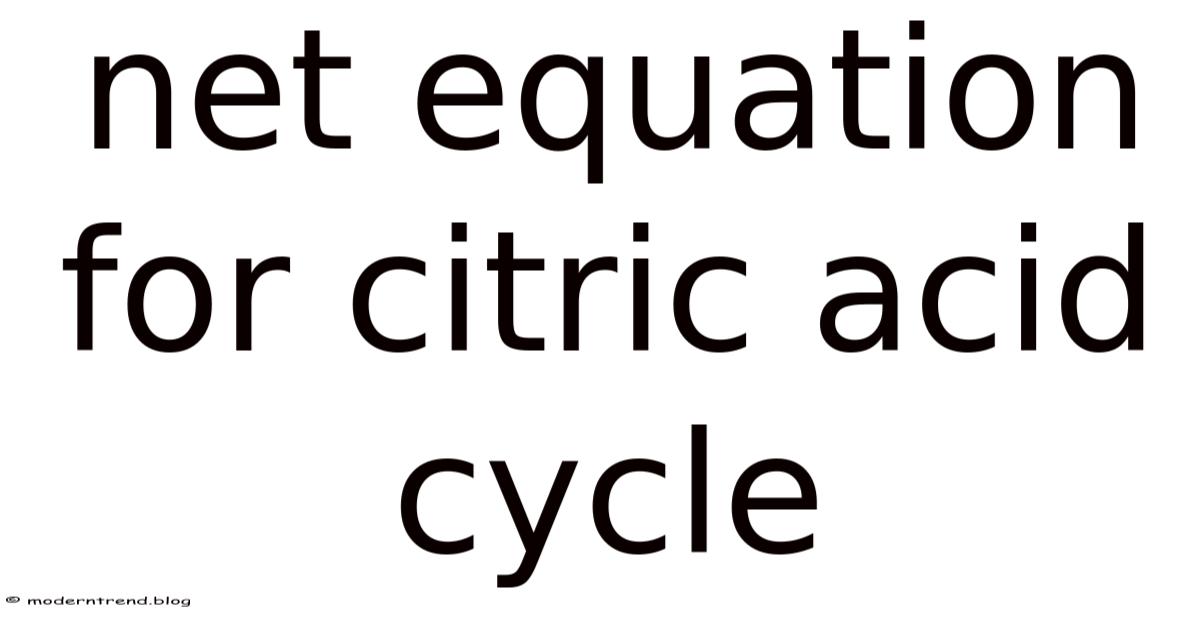
Ever wondered what the overall chemical “balance sheet” looks like for the citric acid cycle?
You’ve probably seen the maze of arrows in textbooks, each step shuffling carbon atoms, NAD⁺, and CoA around like a molecular dance party. But when the dust settles, there’s a single net equation that tells you exactly what goes in and what comes out. Knowing that equation isn’t just academic trivia—it’s the shortcut that lets you see the big picture of cellular respiration in a glance.
What Is the Net Equation for the Citric Acid Cycle?
When we talk about the citric acid cycle (also called the Krebs or TCA cycle), we’re really talking about a series of eight enzyme‑catalyzed reactions that oxidize acetyl‑CoA to CO₂ while harvesting high‑energy carriers. The “net equation” strips away every intermediate—citrate, α‑ketoglutarate, succinate, and the rest—and shows only the starting substrates and final products No workaround needed..
In plain English, the cycle takes one molecule of acetyl‑CoA, combines it with four molecules of water, and uses three molecules of NAD⁺, one molecule of FAD, and one molecule of GDP (or ADP). The result? Two CO₂, three NADH, one FADH₂, one GTP (or ATP), and two CoA‑S‑H that are ready to re‑enter the pathway.
The compact form looks like this:
Acetyl‑CoA + 3 NAD⁺ + FAD + GDP + Pi + 2 H₂O → 2 CO₂ + 3 NADH + FADH₂ + GTP + CoA‑SH
If you prefer the ATP‑centric version, replace GDP + Pi with ADP + Pi → ATP, and you’ll see the same energy yield.
Why It Matters – The Real‑World Reason You Should Care
Understanding the net equation does more than satisfy curiosity. It’s the bridge between biochemistry textbooks and real‑world applications:
- Metabolic engineering: When you tweak a microbe to produce a bio‑fuel, you need to know how many NADH molecules you’re generating per turn of the cycle. That net equation tells you exactly that.
- Medical diagnostics: Elevated lactate or abnormal NAD⁺/NADH ratios often point back to a bottleneck in the TCA cycle. Knowing the stoichiometry helps clinicians interpret lab values.
- Exercise physiology: During intense workouts, muscles rely on the TCA cycle for ATP. The net equation shows why you burn more carbs (acetyl‑CoA) when you’re sprinting versus cruising.
In short, the net equation is the “quick‑look” cheat sheet that lets you predict energy output, carbon loss, and cofactor demand without tracing every arrow.
How It Works – Step‑by‑Step Breakdown
Below is the “inside the kitchen” view of how the cycle turns acetyl‑CoA into the products listed above. I’ll keep the focus on the chemistry that matters for the net equation, skipping the minutiae that most people forget.
### 1. Acetyl‑CoA Enters the Stage
Acetyl‑CoA (a two‑carbon unit attached to coenzyme A) condenses with oxaloacetate (a four‑carbon molecule) to form citrate (six carbons). This step is catalyzed by citrate synthase and consumes no cofactors—just a quick carbon‑shuffle Small thing, real impact..
### 2. Two Oxidations, Two NADH
Citrate is rearranged to isocitrate, then oxidized by isocitrate dehydrogenase. That oxidation releases one CO₂ and reduces NAD⁺ to NADH. The same pattern repeats when α‑ketoglutarate is oxidized to succinyl‑CoA, giving you the second NADH and the second CO₂.
### 3. Substrate‑Level Phosphorylation – GTP (or ATP)
Succinyl‑CoA → succinate is a high‑energy step. The thioester bond in succinyl‑CoA transfers its energy to GDP (or ADP) + Pi, forming GTP (or ATP). This is the only direct ATP‑making move inside the cycle.
### 4. The FAD Step
Succinate is oxidized to fumarate by succinate dehydrogenase, reducing FAD to FADH₂. This is the only flavin‑based reduction in the whole pathway No workaround needed..
### 5. The Final NADH
Fumarate → malate → oxaloacetate completes the circle. Malate dehydrogenase reduces another NAD⁺ to NADH while converting malate to oxaloacetate, ready to accept the next acetyl‑CoA.
### 6. Putting It All Together
If you add up the three NAD⁺ reductions, one FAD reduction, and the substrate‑level phosphorylation, you arrive at the net equation shown earlier. Notice that water appears on the reactant side because two hydration steps (citrate → isocitrate and fumarate → malate) each consume a water molecule.
Common Mistakes – What Most People Get Wrong
-
Leaving out water
Many cheat sheets drop the two H₂O molecules, assuming they’re “obvious.” In reality, those waters are consumed in the hydration steps, and omitting them throws off the atom balance. -
Confusing GTP with ATP
The cycle produces GTP directly, not ATP. Sure, many cells have a nucleoside‑diphosphate kinase that quickly swaps GTP for ATP, but the net equation should reflect the actual product: GTP (or ADP + Pi → ATP if you prefer the latter notation). -
Counting CO₂ twice
Some sources list three CO₂ because they mistakenly add the two released during the oxidative decarboxylations plus the carbon from acetyl‑CoA. The correct total is two CO₂, matching the loss of the two carbons originally in acetyl‑CoA Nothing fancy.. -
Forgetting CoA‑SH
The coenzyme A that entered as acetyl‑CoA is regenerated at the end. Ignoring the CoA‑SH term makes the equation look like CoA disappears into thin air. -
Mixing up NAD⁺ and NADP⁺
The TCA cycle uses NAD⁺, not NADP⁺. The latter is reserved for the pentose phosphate pathway and anabolic reactions. Swapping them changes the redox balance completely.
Practical Tips – What Actually Works When You Need the Net Equation
- Write it out on paper. Start with acetyl‑CoA and list each reaction’s inputs and outputs. Cancel intermediates line‑by‑line; the leftovers are your net equation.
- Balance atoms, then charge. Carbon and oxygen are the easiest; hydrogen and charge follow. If you end up with extra protons, add water or H⁺ as needed.
- Use a spreadsheet. A quick column for each metabolite lets you sum rows and see what cancels. It’s a neat trick for teaching or for checking your work.
- Remember the “four water” rule. Two hydration steps consume water; two dehydration steps (if you count them) release it. In the net equation, you only need the two waters that are truly consumed.
- Swap GTP for ATP only if you’re comparing energy yields. When you calculate ATP equivalents, treat the GTP as one ATP. But keep the original GTP in the biochemical equation.
FAQ
Q: Does the net equation change if the cell uses NAD⁺ vs. NADP⁺?
A: No. The TCA cycle strictly uses NAD⁺. If you see NADP⁺ in a version of the equation, it’s a mistake or a specialized variant found in some bacteria Not complicated — just consistent. Turns out it matters..
Q: How many ATP molecules does one turn of the cycle actually produce?
A: Directly, you get one GTP (≈ 1 ATP). The three NADH and one FADH₂ feed into oxidative phosphorylation, yielding roughly 2.5 ATP per NADH and 1.5 ATP per FADH₂. That adds up to about 10 ATP equivalents per acetyl‑CoA Surprisingly effective..
Q: Can the net equation be written with ADP instead of GDP?
A: Yes, if you prefer the ATP‑centric view. Replace “GDP + Pi → GTP” with “ADP + Pi → ATP” and the rest of the equation stays the same Not complicated — just consistent..
Q: Why is CoA‑SH listed as a product?
A: Acetyl‑CoA donates the acetyl group, but the CoA moiety is regenerated at the end of the cycle. It’s a catalyst, not a consumable, so the net equation must show it returning to the pool Took long enough..
Q: Does the net equation differ in anaerobic organisms?
A: The core stoichiometry stays the same, but some anaerobes run the cycle in reverse (reductive TCA) to fix CO₂, swapping the direction of NADH/FADH₂ production for NADPH/FADH₂ consumption Worth keeping that in mind..
The short version? Plus, the citric acid cycle’s net equation is a tidy snapshot of a complex dance: acetyl‑CoA + 3 NAD⁺ + FAD + GDP + Pi + 2 H₂O → 2 CO₂ + 3 NADH + FADH₂ + GTP + CoA‑SH. Keep it handy, and you’ll always know exactly what the cell is getting out of each turn of the cycle That's the part that actually makes a difference..
And that’s it—no fluff, just the chemistry you need to keep in mind when you’re reading a paper, designing an experiment, or just satisfying that “how does it work?” curiosity. Happy metabolizing!
Beyond the textbook stoichiometry, the net equation serves as a versatile scaffold for a range of practical applications. In metabolic‑flux analysis (MFA), each term is assigned an isotopic label — ^13C‑acetyl‑CoA, ^15N‑glutamate, or ^2H‑water — allowing researchers to trace the fate of carbons through the cycle and quantify pathway activity in vivo. By plugging measured labeling patterns into the balanced equation, one can solve for fluxes that would otherwise be hidden beneath overlapping reactions Not complicated — just consistent..
The equation also highlights points where regulation intersects with energy balance. Take this case: the GTP‑producing step (succinyl‑CoA synthetase) is sensitive to the ADP/ATP ratio, while the NAD⁺‑dependent dehydrogenases are modulated by the NADH/NAD⁺ pool. When a cell shifts from a high‑energy state to a low‑energy state, the net equation predicts a concomitant rise in the demand for ADP and Pi to sustain GTP synthesis, a relationship that can be tested experimentally by measuring changes in nucleotide pools alongside oxygen consumption The details matter here..
In disease contexts, alterations to any term of the net equation have diagnostic significance. Accumulation of succinate or fumarate — detectable as deviations from the expected succinate‑to‑fumarate ratio — points to succinate dehydrogenase inhibition, a hallmark of certain tumors and rare mitochondrial disorders. Likewise, elevated lactate paired with reduced NADH production can signal a shunt away from the cycle toward anaerobic glycolysis, a pattern captured by rewriting the net equation to include lactate dehydrogenase as a side reaction And that's really what it comes down to. Took long enough..
Educators often use the concise format to teach redox bookkeeping. In real terms, by asking students to balance the equation before revealing the answer, they reinforce the principle that every electron removed from a substrate must appear somewhere else — either as reduced NADH/FADH₂ or as released CO₂. The “four water” rule, mentioned earlier, becomes a memorable checkpoint: if the final count of water molecules deviates from two, a mistake in counting hydration/dehydration steps is likely.
Finally, the net equation’s simplicity makes it ideal for quick sanity checks in computational models. When constructing a genome‑scale metabolic reconstruction, the citrate‑cycle subnetwork can be collapsed into a single reaction that mirrors the net equation; this reduces dimensionality while preserving the essential stoichiometric constraints needed for flux balance analysis.
In short, the citric‑acid‑cycle net equation is more than a memorized line — it is a functional tool that links molecular detail to cellular physiology, guides experimental design, and illuminates both normal metabolism and its perturbations. Keep it at hand, and you’ll have a clear, quantitative window into one of life’s most central metabolic cycles.